
DISCUSSION
There are relatively few studies comparing the biomass of the selected understory conifer trees, and few comparisons can be drawn. The only directly comparable equations are those from Brown (1978); total aboveground biomass as a function of height was presented for dominant ponderosa pine, grand fir (Abies grandis (Doug!.) Forbes), and lodgepole pine in the Rocky Mountains. Grand fir is closely related to white fir and the two are difficult to distinguish in Oregon. Graphical comparison (Figures 2, 3, 4) shows the biomass of the Rocky Mountain trees to be intermediate between “open” and “dense” grown southern Cascade trees. The equations may thus represent a broader range of habitats than the sample locations indicate. The most comprehensive set of biomass equations for Pacific Northwest plants {Gholz et al., 1979) uses pooled data sets or data from areas outside Oregon and Washington, but note that care must be taken to evalate the applicability of an equation to a specific site separated from the sample site. The caution also applies to the equations ln this study.
Biomass of trees is often estimated using sample trees that are
open-grown dominants. Using Brown’s (1978) data for small trees grown in shaded environments would overestimate total biomass by a factor of 2 to 5 (Figure 2). Knowledge of the specific microsite as well as the general habitat will enable more precise biomass prediction.
The biomass equations show generally decreasing goodness-of-fit as fuel size classes become larger. Over the same series of size classes reported here, Brown (1976) found coefficients of determination for biomass and branch diameter regressions to decrease from 0.93 to 0.82, for eleven western conifers including lodgepole and ponderosa pine. In both studies there were fewer observations in the larger size classes. The shade tolerant species in this study (white fir, Shasta red fir, mountain hemlock) tended to have higher foliar biomass than shade intolerant species (lodgepole pine, ponderosa pine), which is also true of most of the species that Brown (1976) studied, except ponderosa pine.
Prediction of age from size characteristics is difficult, even for
the relatively small ranges in size included in this study. Oliver (1981) has reviewed the relation of tree age to stand architecture and concluded that a narrow range of tree ages can display the vertical and diameter distribution normally associated with all-age forests. Conversely, stems of similar height and diameter may represent significantly different ages.
Relative shade tolerance of species can be estimated by comparing the foliar biomass of trees grown in shaded and open environments. The average foliar biomass by species in dense overstory and open overstory conditions, and the ratio of dense to open foliar biomass, is shown in Table 10. Mountain hemlock has a ratio approaching 1 .0, suggesting it is able to maintain leaf area quite well in shade. Shasta red fir has a much lower ratio, followed by white fir, ponderosa pine, and lodgepole pine. This ranking differs slightly from the rankings of Hinore (1979) who reviewed autecological characteristics of northwestern tree species (Table 10, right column). The rankings of the two fir species and the two pine species are reversed.
The ratio of Shasta red fir is almost twice that of white fir, yet from Hinore’s ranking it is less shade tolerant and should maintain relatively less leaf area in the shade than in the open. The successional patterns of Shasta red fir are not wel I documented. It has been described as climax in some areas and seral to white fir or to mountain hemlock elsewhere (Frankl in and Dyrness, 1973). In the locations at Crater Lake where it was sampled.
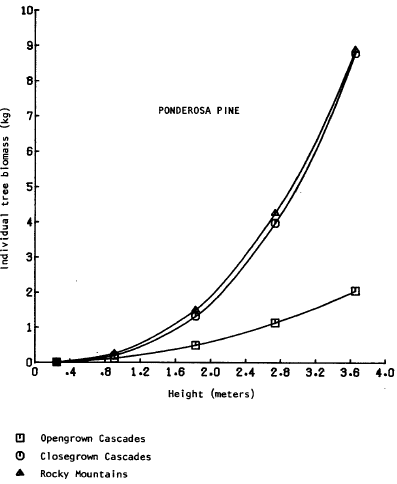
Figure 2. Tree biomass vs. height – ponderosa pine.
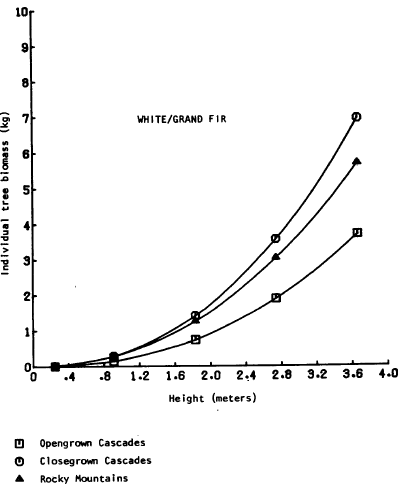
Figure 3. Tree biomass vs. height – white/grand fir.
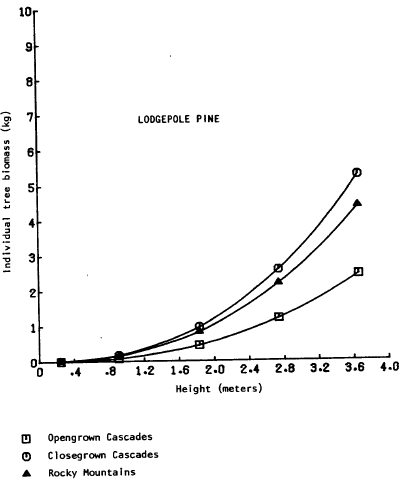
Figure 4. Tree biomass vs. height – lodgepole pine.
Shasta red fir appears to have moderately high shade tolerance based on its ability to maintain leaf area under shade. This also suggests, due to a partial association of shade tolerance and climax status, that Shasta red fir may be a climax dominant in this area.
The ratio reversal of ponderosa and lodgepole pine foliar biomass is probably due to the specific sampling locations. The lodgepole pine samples were collected in a stand where it is seral to mountain hemlock, while the ponderosa pine samples were collected in a climax stand of ponderosa pine. For similar basal areas where overstories were dense, the hemlock stand appeared to provide much more shade, reducing the foliar biomass of shaded understory lodgepole pine and also the foliar biomass ratio. Had ponderosa pine samples been collected in stands where white fir is climax, a similar reduction in foliar biomass of the pine might have occurred. The ratios of ponderosa pine and lodgepole pine are, therefore, not directly comparable, and the foliar biomass ratio reversal is not of major significance.
The white fir foliar biomass ratio is only slightly above that of ponderosa pine when both are sampled from areas where they are climax dominants. This suggests that white fir, commonly observed to grow in doghair thickets in mixed conifer forest understories, may be less shade tolerant than it often is assumed to be. In Montana, where grand fir appears to have an ecological role similar to white fir, most understory trees from 1-4 m tall are between 40-120 years old (Antos and Habeck, 1981). They are able to survive, but grow very slowly; new establishment is limited to openings in the stand. Similar age structures were found in this study (Table 2). Much of the white fir reproduction seems to have originated in a period of several decades after fire suppression became effective and stands were relatively open. Since the time of understory closure, light and moisture competition have all but eliminated further tree establishment.
Other pages in this section