
Poor growing conditions are reflected in the small size of the trees. Average diameter is less than 15 cm and average height less than 10 m. Maximum values for each parameter are only twice these low averages. Density of all live trees is 800 per hectare (325/ac) illustrating the open grown nature of the stand. Dwarf mistletoe was common on trees exceeding 100 years of age, and absent on younger trees. However, some of the 100-year-old infected trees were only 1 m tall.
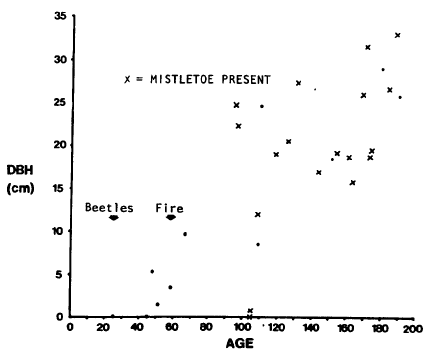
| FIGURE 2. Age-diameter relationship for a climax P. contortastand. |
Although the age-diameter relationship is positive, there is a wide range of ages for any given diameter. While not shown graphically here, significant overlap in age-height relationships also occurs. Predicting structure from diameter or height structure is not feasible. Inference about age structure depends on rather complete age analysis.
The age class analysis on the plot revealed a distinct three-aged stand (Figure 3). There is a maximum peak at 160-180 years, followed by a significant decline in the younger 140-160 and 120-140 age groups. A second peak occurs at 100-120 years, followed by a similar decline in the next two younger age classes. A third peak occurs in the 40-60 year old class, followed by a decline in the 20-40 year age class. The apparent peak in the 0-20 year class is the result of continuing regeneration which usually succumbs; the two trees in this class are 10 cm tall and a third tree of similar size died on the plot in 1979.
This age structure is quite unusual and appears consistent with the fire-insect-fungi hypothesis of Geiszler et al. (1980). In this stand, three age classes are present rather than the two hypothesized, but assuming a 60-year “cycle” and a 200-year time frame, the age structure is consistent.
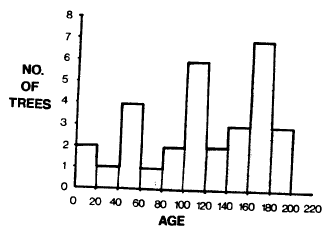
| FIGURE 3. Age structure of the P. contorta stand. |
Validation of the processes involved was partially possible. In a limited search of the area, no evidence for disturbance in the 100-120 or 160-180 year period was uncovered; little time was devoted to this, however. A fire scar sampled from a live tree indicated the last fire was in 1922. Some thirty years later, in 1954-56, bark beetles killed some of these scarred trees, as evidenced by examination of the age of callus tissue on dead and down logs that had fire scars. Over the last 25 years, these dead trees have fallen over the area and provided the fuel for the 1980 fire.
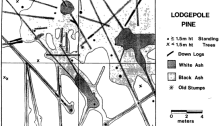
The map of fire spread over the plot (Figure 4) indicates that these logs were the primary corridors of fire spread. Where stand density was greatest, some surface fire spread occurred, but where the stand was open (i.e., lower two quadrats) logs were the only fuel burning. In the relatively denser portions of the plots, there was enough needlefall. At the time they burned, most were Class 2-3 logs, hard on the exterior but punky and moist inside. The fire moved by conduction, drying out the logs ahead of the very slow flaming or smoldering front. It is thought that the spread on the plot occurred over a period of several days.
This slow moving fire created matrix of burned and unburned patterns on the plot. It helps to explain the existence of fire scars a meter or more long on the trunks of older trees. These older scars may have been created by heat radiation from a burning log near one side of the tree; the base of the tree may not have been burned at all, but the cambium could be killed through the thin bark. This theory also explains why only some of the trees are scarred; those not near logs have a high probability of remaining unscarred.
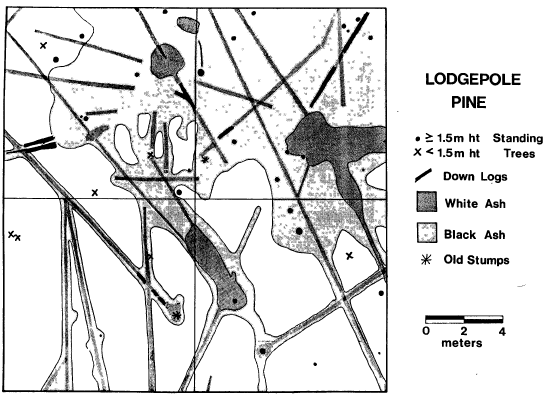
| FIGURE 4. Map of standing trees, down logs, and black/white ash areas onP. contorta plot. |
This type of fire spread does not easily conform to existing fire behavior models. Where the litter was continuous, Albini’s fuel model 8 (closed litter) appeared to closely approximate the low intensity of the fire. For the range of fire weather conditions during the week of the fire (Table 2), rate of spread ranged from 0 to 0.66 m/min, and fireline intensity ranged from less than I to 5 Kcal-sec-m. Flame length was predicted to range from 0 to 0.3 m, which under the temperature and wind conditions resulted in a predicted 0-1 m scorch height. This was generally true for the plot, although a few trees with very low hanging mistletoe infections ignited and resulted in much higher scorch heights.
| TABLE 2. Fire weather in late June-early July 1980 during the Sharp Peak prescribed fire. | |
| ShadeDry Bulb Temp, FRelative Humidity
I H TL FM, % 10 H TL FM, % 100 H TL FM, % Live Fuel Moisture Midflame Windspeed Slope |
055-7530-50
8-11 8-10 17-19 150 1-5 O |
Both stand mortality and regeneration will be followed over time on this plot. The fire did kill some trees and weakened others; there was some bark beetle activity on the plot September 1980. The location of subsequent regeneration in relation to black and white ash corridors will be mapped as it occurs.
Discussion
The climax lodgepole pine stands at Crater Lake appear to have a similar structure to the ones studied by Geiszler et al. (1980) nearby. Although evidence is not complete on the process involved in the generation of multi-aged stands such as these, the evidence to date is consistent with the fire-fungi hypothesis. Fire frequency appears to have been historically consistent in these stands: about I fire every 60 years. Whether this is controlled by climate or controlled by fuel accretion after beetle outbreaks requires further investigation. Fire intensity appears to have been low to moderate in the past, and in most stands the influence of 60 years of fire suppression has been very minimal.
A policy of allowing natural fires to burn in these climax lodgepole stands is quite consistent with the objective to maintain natural ecosystems in this area. However, in those areas where lodgepole pine is a seral member of the community, other fire strategies are appropriate. Prescribed fire may be needed in the Abies concolor zone to reduce fuel loads and understory density, and fire suppression may be quite appropriate in the Tsuga-Abies forest zone (Zobel and Zeigler 1978) due to an overabundance of man-induced fire in the late 1800s.
It is hoped that further investigation into the stand dynamics of the climax lodgepole pine forests at Crater Lake and testing of the fire insect-fungi hypothesis will be supported by National Science Foundation grant money. A decision on the proposal should be known by late Spring 1981.
Literature Cited
Albini, F. 1976. Estimating Wildfire Behavior and Effects. USDA For. Serv. Gen. Tech. Rpt. INT-30. Intmt. For. and Range Exp. Sta., Ogden, UT. 92 pp.
Burgan, R. 1979. Fire Danger/Fire Behavior Computations with the Texas Instruments TI-59 Calculator: User’s Manual. USDA Forest Service General Technical Report INT-61. Intmt. For. and Range Exp. Sta., Ogden, UT. 21 pp.
Franklin, J.F. and C.T. Dyrness. 1973. Natural Vegetation of Oregon and Washington. USDA For. Serv. Gen. Tech Rpt. PNW-8. Pac. NW For. and Range Exp. Sta., Portland, OR. 417 pp.
Geiszler, D.R., R.I. Gara, C.H. Driver, V.F. Gallucci, and R.E. Martin. 1980. Fire, fungi, and beetle influences on a lodgepole pine ecosystem of south-central Oregon. Oecologia (Berl.) 46:239-243.
McNeil, R.C. and D.B. Zobel. 1980. Vegetation and fire history of a ponderosa pine-white fir forest in Crater Lake National Park. Northwest Science 54(1):30-46.
Zeigler, R. 1978. The vegetation dynamics of Pinus contorta forest, Crater Lake National Park. M.S. thesis, Oregon State Univ. 182 pp.
Zobel, D.B. and R. Zeigler. 1978. Vegetation Dynamics of Lodgepole Pine Forest, Crater Lake National Park. Final Report, Contract CX-9000-6-0064. 32 pp.
Other pages in this section
*** previous title *** --- *** next title ***